Превращения моно и дисахаридов
Дыхание это экзотермический процесс ферментативного окисления моносахаридов до воды и диоксида углерода:
Дыхание является важнейшим источником энергии для человека. Для осуществления процесса дыхания необходимо большое количество кислорода.
При недостатке кислорода или его отсутствии происходит процесс брожения моносахаридов. Существует несколько видов брожения, в которых принимают участие различные микроорганизмы.
Спиртовое брожение осуществляется при участии ферментов дрожжей по следующей схеме:
В результате реакции спиртового брожения, под действием комплекса ферментов дрожжей, образуется две молекулы этилового спирта и две молекулы диоксида углерода. Моносахариды сбраживаются дрожжами с различной скоростью. Наиболее легко сбраживается глюкоза и фруктоза, труднее манноза, практически не сбраживается галактоза – основной углевод молока. Пентозы дрожжами не сбраживаются. Наряду с моносахаридами глюкоза и фруктоза, дрожжи могут сбраживать дисахариды мальтоза исахароза, так как дрожжи обладают ферментами, способными разложить молекулы этих двух дисахаридов до глюкозы и фруктозы (ά -гликозидаза и β-фруктофуранозидаза). Спиртовое брожение играет важную роль в процессе производства пива, спирта, вина, кваса, в хлебопечении. Наряду с главными продуктами брожения – этиловым спиртом и диоксидом углерода, при спиртовом брожении образуются побочные и вторичные продукты брожения: глицерин, уксусный альдегид, уксусная кислота, изоамиловый и другие высшие спирты. Эти продукты влияют на органолептические свойства продуктов, часто ухудшают их качество.
Молочнокислое брожение осуществляется при участии ферментов молочнокислых бактерий:
В результате реакции молочнокислого брожения под действием комплекса ферментов образуется две молекулы молочной кислоты. Молочнокислое брожение играет важную роль в процессе производства кисломолочных продуктов, кваса, квашении капусты.
Маслянокислое брожение осуществляется при участии ферментов маслянокислых бактерий:
В результате реакции маслянокислого брожения образуется молекула масляной кислоты две молекулы диоксида углерода и водород. Этот процесс происходит на дне болот при разложении растительных остатков, а также при возникновении инфекции маслянокислыми микроорганизмами в процессе производства продуктов питания.
Лимоннокислое брожение осуществляется при участии ферментов плесневого гриба Aspergillus niger:
В результате реакции лимоннокислого брожения образуется молекула лимонной кислоты. В основе этой реакции лежит процесс получения лимонной кислоты.
Карамелизация. Реакция карамелизации осуществляется при нагреве свыше 100 °С растворов глюкозы, фруктозы, сахарозы. При этом происходят различные превращения углеводов. При нагревании сахарозы в слабокислой среде происходит частичный гидролиз (инверсия) с образованием глюкозы и фруктозы. От молекул глюкозы и фруктозы при нагревании может отщепляться три молекулы воды, происходит дегидратация с образованием оксиметилфурфурола, дальнейшее разрушение которого приводит к разрушению углеродного скелета и образованию муравьиной и левулиновой кислот. Оксиметилфурфурол образуется при нагревании растворов углеводов низкой концентрации – 10 — 30 %, это вещество имеет коричневый цвет и специфический запах пропеченной корочки хлеба.
На первом этапе реакции карамелизации от молекулы сахарозы отщепляется две молекулы воды. Образуется карамелан, состоящий из ангидроколец, содержащих в кольце двойные связи (дигидрофуран, циклогексанолон и другие соединения), которые имеют коричневый цвет. На втором этапе отщепляется три молекулы воды и образуется карамелен, имеющий темнокоричневый цвет. На третьем этапе происходит конденсация молекул сахарозы и образуется карамелин, имеющий темнокоричневый цвет, плохо растворимый в воде. Карамелизация сахарозы осуществляется при содержании сахарозы 70 — 80 %.
Схема превращения дисахаридов при нагревании:
ДИСАХАРИДЫ → КАРАМЕЛАН → КАРАМЕЛЕН → КАРАМЕЛИН
Схема превращения моносахаридов при нагревании:
ГЛЮКОЗА, ФРУКТОЗА → АНГИДРИДЫ → ОКСИМЕТИЛФУРФУРОЛ
Меланоидинообразование или реакция Майяра. Реакция взаимодействия восстанавливающих дисахаридов и моносахаридов с аминокислотами, пептидами, белками. В результате взаимодействия карбонильной (альдегидной или кетонной) группы углеводов и аминогруппы белков и аминокислот происходят многостадийные превращения продуктов реакции с образованием глюкозоамина, который подвергается перегруппировке по Амадори и Хейтсу, затем образуются меланоидиновые пигменты, имеющие темно-коричневый цвет, специфический вкус и запах. Реакция меланоидинообразования является основной причиной неферментативного потемнения пищевых продуктов. Такое потемнение происходит при выпечке хлеба, при сушке солода, при кипячении сусла с хмелем в производстве пива, при сушке фруктов. Скорость реакции зависит от состава взаимодействующих продуктов, рН среды, температуры, влажности. В результате реакции меланоидинообразования снижается содержание углеводов и аминокислот, в том числе и незаманимых, на 25 %, что приводит и к изменению качества готового продукта, снижению его пищевой и энергетической ценности. Имеются сведения, что продукты реакции меланоидинообразования обладают андиоксидантными свойствами, снижают усвоение белков.
Схема взаимодействия восстанавливающих дисахаридов и моносахаридов с аминокислотами в упрощенном виде:
УГЛЕВОД + АМИНОКИСЛОТА → ГЛЮКОЗОАМИН →
Понравилась статья? Добавь ее в закладку (CTRL+D) и не забудь поделиться с друзьями:
Источник
Метаболизм фруктозы и галактозы
Метаболизм фруктозы и галактозы включает пути использования их для синтеза других веществ (гетерополисахаридов, лактозы и др.) и участие в энергообеспечении организма. В последнем случае фруктоза и галактоза превращаются в печени либо в глюкозу, либо в промежуточные продукты её метаболизма. Таким образом, в результате фруктоза и галактоза наряду с глюкозой могут быть окислены до СО2 и Н2О или использованы на синтез гликогена и триацилглицеролов.
Причиной нарушения метаболизма фруктозы и галактозы может быть дефект ферментов, катализирующих промежуточные реакции их обмена. Эти нарушения встречаются относительно редко, но могут представлять достаточно серьёзную опасность, так как накапливаемые промежуточные метаболиты фруктозы и галактозы обладают токсичностью.
А. Метаболизм фруктозы
Значительное количество фруктозы, образующееся при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Другая часть фруктозы всасывается с помощью белка-переносчика, т.е. путём облегчённой диффузии.
Метаболизм фруктозы (рис. 7-69) начинается с реакции фосфорилирования (реакция 1), катализируемой фруктокиназой с образованием фруктозо-1-фосфата. Фермент обнаружен в печени, а также в почках и кишечнике. Этот фермент обладает абсолютной специфичностью, поэтому, в отличие от глкжокиназы, инсулин не влияет на его активность. Последнее обстоятельство объясняет, почему уровень выведения фруктозы в моче у больных сахарным диабетом и здоровых не отличается. Фруктозе-1-фосфат не может превращаться во фруктозо-6-фосфат из-за отсутствия соответствующего фермента. Вместо этого фруктозо-1-фосфат далее расщепляется фруктозо-1-фосфатальдолазой (альдолаза В) на глицеральдегид и дигидроксиацетон-3-фосфат (реакция 2). Последний является промежуточным продуктом гликолиза и образуется в ходе реакции, катализируемой фруктозо-1,6-бисфосфосфатальдолазой (альдолаза А). Глицеральдегид может включаться в гликолиз после его фосфорилирования с участием АТФ (реакция 3). Две молекулы триозофосфатов либо распадаются по гликолитическому пути, либо конденсируются с образованием фруктозо-1,6-бисфосфата и далее участвуют в глюконеогенезе (реакции 8, 7, 5, 9). Фруктоза в печени включается главным образом во второй путь. Часть дигидроксиацетон-3-фосфата может восстанавливаться до глицерол-3-фосфата и участвовать в синтезе триацилглицеролов.
Следует отметить, что включение фруктозы в метаболизм через фруктозо-1-фосфат минует стадию, катализируемую фосфофруктокиназой (реакция 6), которая является пунктом метаболитического

Рис. 7-69. Метаболизм фруктозы. а — превращение фруктозы в дигидроксиацетон-3-фосфат и глицеральдегид-3-фосфат; б — путь включения фруктозы в гликолиз и глюконеогенез; в — путь включения фруктозы в синтез гликогена.
контроля скорости катаболизма глюкозы. Этим обстоятельством можно объяснить, почему увеличение количества фруктозы ускоряет в печени процессы, ведущие к синтезу жирных кислот, а также их этерификацию с образованием триацилглицеролов.
Источник
Пути вхождения углеводов в гликолиз. Полное окисление фруктозы (начиная с фруктокиназной реакции). Биоэнергетика процесса, фруктозурия.
Полное окисление фруктозы. Установлено, что фруктоза, присутствующая в свободном виде во многих фруктах и образующаяся в тонкой кишке из сахарозы, всасываясь в тканях, может подвергаться фосфорилированию во фруктозо-6-фосфат при участии фермента гексокиназы и АТФ:
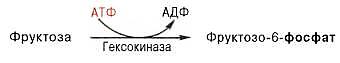
Эта реакция ингибируется глюкозой. Образовавшийся фруктозо-6-фос-фат либо превращается в глюкозу через стадии образования глюкозо-6-фосфата и последующего отщепления фосфорной кислоты, либо подвергается дальнейшим превращениям. Из фруктозо-6-фосфата под влиянием 6-фосфофруктокиназы и АТФ образуется фруктозо-1,6-бисфос-фат:

Далее фруктозо-1,6-бисфосфат может подвергаться дальнейшим превращением по пути гликолиза. Таков главный путь включения фруктозы в метаболизм мышечной ткани, почек, жировой ткани.
В печени, однако, для этого существует другой путь. В ней имеется фермент фруктокиназа, который катализирует фосфорилирование фруктозы не по 6-му, а по 1-му атому углерода:

Эта реакция не блокируется глюкозой. Образовавшийся фруктозо-1-фосфат расщепляется затем под действием кетозо-1-фосфатальдолазы на диоксиацетонфосфат и D-глицеральдегид:
Фруктозо-1-фосфат Диоксиацетонфосфат + D-глицеральдегид.
Образовавшийся D-глицеральдегид под влиянием соответствующей киназы (триокиназы) подвергается фосфорилированию до глицеральде-гид-3-фосфата. В этот же промежуточный продукт гликолиза переходит и дигидроксиацетонфосфат.
Существует врожденная аномалия обмена фруктозы, или эссенциальная фруктозурия, которая связана с врожденным недостатком фермента фрук-токиназы, т.е. в организме не образуется фруктозо-1-фосфат. В результате обмен фруктозы возможен только путем фосфорилирования до фрук-тозо-6-фосфата, но эта реакция тормозится глюкозой, вследствие чего фруктоза накапливается в крови. «Почечный порог» для фруктозы очень низок, поэтому фруктозурия обнаруживается уже при концентрации фруктозы в крови 0,73 ммоль/л.
Галактоза. Основным источником галактозы является лактоза пищи, которая в пищеварительном тракте расщепляется до галактозы и глюкозы. Обмен галактозы начинается с превращения ее в галактозо-1-фосфат. Эта реакция катализируется галактокиназой с участием АТФ:


В следующей реакции в присутствии УДФ-глюкозы фермент гексозо-1-фосфатуридилилтрансфераза катализирует превращение галактозо-1-фосфата в глюкозо-1-фосфат, одновременно образуется уридиндифосфат-галактоза (УДФ-галактоза):
Образовавшийся глюкозо-1-фосфат в дальнейшем либо переходит в глюкозо-6-фосфат и далее подвергается уже известным превращениям, либо под влиянием фосфатазы образует свободную глюкозу, а УДФ-га-лактоза подвергается весьма своеобразной эпимеризации:

Затем УДФ-глюкоза-пирофосфорилаза катализирует расщепление УДФ-глюкозы с образованием глюкозо-1-фосфата:

Понравилась статья? Добавь ее в закладку (CTRL+D) и не забудь поделиться с друзьями:
Источник